Exogenous feed enzymes have been in commercial use in swine diets for almost 30 years. Although proteases represent a small proportion of total sales compared to carbohydrases and phytases, their application in swine diets has interest, particularly on the utilisation of amino acids (AA) in plant-based protein sources and reducing the detrimental effects of nitrogen (N) excretion on the environment. However, supplementation of proteases into pig diets has resulted in positive, inconclusive, inconsistent or, in some cases, depressive effects on performance.
The first positive reported effects from protease supplementation are from the 1950s when in a series of experiments with piglets weaned at 6-10 days of age, Lewis et al. (1955) fed nutrient-adequate diets with the addition of pancreatin, pepsin, pepsin plus pancreatin, fungal protease, papain, or a proteolytic enzyme derived from Aspergillus oryzae and noted an increase in relative gain by 19, 24, 29, 0, 24 or 29% respectively. Other more recent studies have also demonstrated improvements in pig performance with protease supplementation. For example, in weanling pigs, protease supplementation improved growth rate and N digestibility (Tactacan et al., 2016). Wang et al. (2011) and Zuo et al. (2015) also reported an increase in growth performance of weanling pigs with the latter reporting increased protein digestibility. Supplementing finishing pig diets with a commercial protease produced by Bacillus licheniformis for 9 weeks improved gain to feed ratio and protein digestibility (Lei et al., 2017) which is in line with previous work (Cowieson and Roos, 2016; Upadhaya et al., 2016). The inclusion of an exogenous protease may increase the rate or extent of hydrolysis of dietary protein, thereby making more peptides and AA available for utilization in the small intestine (Guggenbuhl et al., 2012; Yu et al., 2016). However, in some studies no beneficial effects of addition of protease on growth performance were observed (O’Doherty and Forde, 1999; Thacker 2005; Zamora et al., 2011; Guggenbuhl et al., 2012; Mc Alpine et al., 2012; Chen et al., 2017) even though improvements in protein and AA digestibility were observed (Guggenbuhl et al., 2012; Chen et al., 2017) (Table 1). The lack of correlation between digestibility and performance effects is not uncommon and should not be surprising if intake is simultaneously depressed (Walk et al., 2018).

Table 1. Influence of protease supplementation in corn- or sorghum-based diets on growth and apparent ileal digestibility (AID, %) when fed to pigs from 18 to 44 kg (modified from Chen et al., 2017)
| Diet | Protease | ADG, g | ADFI, g | G:F | AID CP | AID GE |
|---|---|---|---|---|---|---|
| Corn | - | 760 | 1375 | 0.555 | 81.7 | 84.5 |
| + | 752 | 1372 | 0.551 | 85.7 | 87.2 | |
| Sorghum | - | 770 | 1443 | 0.535 | 81.8 | 85.5 |
| + | 803 | 1502 | 0.539 | 86.2 | 86.8 | |
| Diet | 0.02 | 0.001 | 0.03 | 0.87 | 0.78 | |
| Protease | 0.33 | 0.31 | 0.99 | 0.01 | 0.06 | |
| Interaction | 0.10 | 0.25 | 0.57 | 0.90 | 0.53 |
Inconclusive or depressive results by feeding protease have also been reported that should be taken in consideration. For example, O’Shea et al. (2014) evaluated the use of protease in finisher pigs fed wheat- and barley-based diets with 21% rapeseed meal and 30% wheat dried distillers grains with solubles. Protease supplementation significantly increased apparent ileal N digestibility but significantly decreased apparent total tract N digestibility and overall feed intake (FI) and final body weight (BW). There was no effect of protease on urine or faecal N. The authors hypothesised the improvement in N digestibility may be indicative of an increased AA uptake in the small intestine and a surplus of free AA which may have resulted in a reduction in FI via an aminostatic feedback mechanism. However, a dietary surplus of N or free AA is typically excreted in the urine or faeces (Monteiro et al., 2010) and this was not the case. Therefore, the improvement in ileal N digestibility in this experiment is most likely the result of depressed intake and thus possibly reduced transit rate, rather than absorption or re-partitioning of surplus AA or N in the body. This resulted in a reduction in ileal digestible AA intake on a daily basis, despite improved AA digestibility, which resulted in a significant decrease in final BW. It is for that reason that digestibility data in the absence of intake data are of limited value.
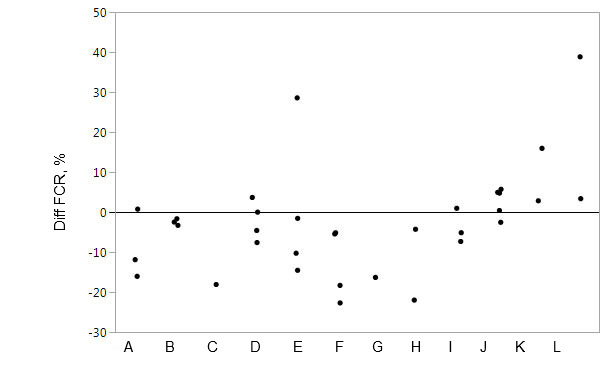
Divergent results may be caused by differences in the period and length of supplementation, in the composition of the experimental diets, in differences across the nature of proteases evaluated, in the supplementation level or in the concurrent supplementation with other enzymes. Combination with other enzymes such as phytase in high doses and carbohydrases can improve the quality of the feed by absorbing part of the potential effect of exogenous proteases (dos Santos et al., 2017), leading to such inconsistency (Lee et al., 2018). Protease supplementation should theoretically improve the nutritive value of dietary protein sources in pig diets when the animal does not produce enough protease of its own but theory conflicts with results reported in the scientific literature. This range in responses between experiments and protease sources is large (Figure 1 & 2; Lee et al., 2018). Further work is required to identify the conditions under which specific proteases will consistently bring benefits in pig production with regards to AA and N utilisation and excretion but in particular growth rate and/or efficiency.