

It represents a coordinated system in which the physical barrier, composed by epithelium, the resident immune cell, the local microenvironment, and its microbiome, work together to maintain the body homeostasis.
Skin is composed by three main layers and by adnexa including hair follicles and sebaceous and sweat glands (figure 1):

- The epidermis (20-26 μm thick), first external barrier, where, continuous desquamation of keratinocytes, desiccation, low pH, sebum and fatty acids and commensal bacteria (up to 1012microorganism/m2), guarantee the integrity and make difficult for pathogens to penetrate it;
- The dermis composed by few fibroblasts, collagen and elastin fibers, contains blood, lymphatic vessels and cells of the innate and acquired immunity;
- The subcutis, composed by a fatty subepidermal layer, with some blood and lymphatic vessels.
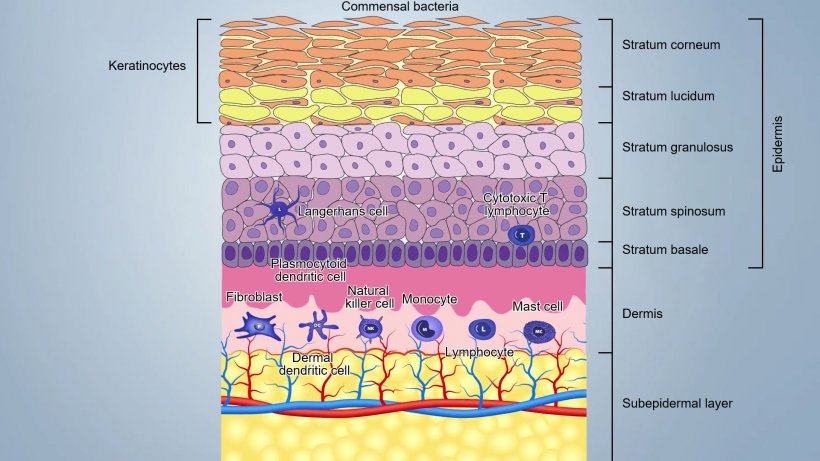
These layers are tightly attached and form a barrier that varies from 50 to 120 μm, depending on the location of the body: in the groin and axillae they are thinner than the other parts of the body.
Immunological components of the epidermis
Keratinocytes are the first defensive cells, expressing a wide range of PRRs (Pathogen Recognition Receptors) like TLR (TLR-1, TLR-2, TLR-4, TLR-5 and TLR-6) and c-type lectins; they also secrete a wide range of pro-inflammatory cytokines and antimicrobial peptides (AMPs). Indeed, keratinocytes can recognize PAMPs (Pathogen Molecular Pattern) by PPRs and trigger inflammation secreting interleukins (IL-1), IFNs, cytokines, growth factors , chemokines, defensins. They can also express MHCII and present antigen.
The antigen trapped on the surface is captured and internalized by Langerhans cells (LC), a special subset of epidermal dendritic cells located between keratinocytes of the basal layer and the stratum spinosum. LCs process antigen and present it to the nearby helper T cells. In porcine epidermis, LCs have a typical dendritic cell (DC) morphology and express MHCII molecules and surface molecules such as CD1, CD172 and CD207.
Epidermal LC are also necessary to sustain an effective cytotoxic response and for Th17 differentiation triggered by secretion of IL-6, IL-1β and IL-23. In pig, 50-70% of the dendritic cells of the skin are LC.
Porcine epidermis also contains a small number of γδT cells, most of them are skin resident - long term cytotoxic memory T cells. Some of these cells, when activated, can kill infected keratinocytes directly and recruit other cells.
Immunoglobulins can cross the skin by transudation and IgG1, IgG2 and IgA can be also locally synthesized.
Video 1. Immune response in the skin.
Immunological components of the dermis
CD1+ dendritic cells represent the dominating and most important immune cells of the dermis. A subset of these cells (dermal dendritic cells) can present the antigen and induce primary T-cell response in the lymph node, while another subset (plasmocytoid dendritic cells) is specialized in the production of a very large quantity of interferon type I in response to recognition of viral nucleic acids.
The dermis contains a large number of lymphocytes which has been estimated to even exceed the number of cells in the blood circulation. T-cell are addressed to dermis by the cutaneous leukocyte antigen (CLA). The major subsets of T-cells are CD4+ Th1, Th2 and Th17.
A subset of circulating T cells, that home to skin and produce IL-22, have been identified and are able to maintain barrier function on exposed body surfaces.
Macrophages (CD14+) are rare in the dermis but monocytes, which rapidly differentiate into macrophages, are efficiently recruited after injury and inflammation.
In dermis there are Natural killers cells (NK) as well; they eliminate virally infected cells and cancer cells with a not MHC-restricted cytotoxicity. NKs are activated by interferons or other cytokines released by macrophages or keratinocytes. NK cells express inhibiting receptors (KIR) that recognise MHC–I and avoid undesirable attacks on self. They can kill target cells through perforin–granzyme pathway.
Mast cells are also important cells involved in inflammation in response to pathogens. MCs serve as sentinel cells being able to recognize danger signals via TLRs and quickly release inflammatory mediators such as histamine, leukotrienes, prostaglandin, pro-inflammatory and immunemodulator cytokines (IL-1, IL-3 , Il-5, IL-6, GM-CSF, IL-4, IL-10, IL-13) and many chemokines. This activity can be induced by different inflammatory stimuli both IgE and non-IgE dependent. The mast cell can also function as antigen presenting cells MHC II-restricted and express specific molecules (B7-1, B7-2, CD40) for activation of T lymphocytes.
Parasitic and bacterial products can be internalized and processed by mast cells and then presented to T lymphocytes.
More specifically, during type I hypersensitivity reactions (Allergy), mast cells are involved in the inflammatory response, releasing mediators when allergen specific IgE, linked to high-affinity immunoglobulin E receptors (FcεRI) on their cell surface, react to a second exposition to the same allergen.
