

The main role of the respiratory system is to capture oxygen from the air as well as to eliminate CO2 from tissue catabolism. In the pig, it also plays an important role in the thermoregulation process, as the pig does not have developed sweat glands. Excess heat is eliminated through an evaporative system called thermal polypnea.
The first part of the respiratory system, the conductive zone or the respiratory zone, runs from the nasal cavity to the last branches of the bronchioles, and also includes the nasal and paranasal sinuses which are connected to the nasal cavity, the nasopharynx and the larynx. All of these tubular structures are lined with ciliated pseudostratified epithelium with goblet cells (Figure 1), which is responsible for one of the main defense systems of the respiratory mucosa, the mucociliary system. This system is made up of the cilia of the respiratory epithelium cells, along with secretions from the goblet cells and the seromucous glands that are below the respiratory mucosa. Their main function is to eliminate particles that enter via the inhaled air. The glands secrete a clear, low viscosity, protein-rich liquid between the cilia, forming a medium that promotes the upward movement of the cilia. The mucus secreted by the goblet cells is deposited on the cilia and allows the inhaled particles to adhere. The upward movement of the cilia pushes the mucus toward the upper respiratory tract; the mucus can then be swallowed and pass into the digestive tract, where it is digested, or be expelled out through the mouth and/or nasal cavity. Agents such as Mycoplasma hyopneumoniae, which causes the loss of cilia, or viruses such as swine flu or respiratory coronavirus, which cause the destruction of epithelial cells, carry out their pathogenic action by destroying this defense system.

Associated with the respiratory mucosa are broncus-associated lymphoid tissues (BALT) where the interaction between lymphocytes and the antigens from external pathogens takes place, causing a specific defensive response, thus avoiding a systemic immune response. This is characterized by a prevalence of humoral immune response through IgA, which is selectively secreted on the surface of the mucosa by means of active transport.
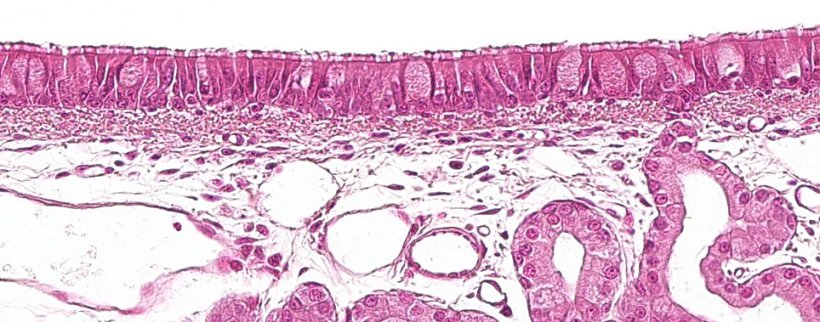
Figure 1: Ciliated pseudostratified epithelium with goblet cells, characteristic of the respiratory system.
Following the respiratory bronchioles comes the respiratory portion, made up of the alveoli, which are defined by a thin wall, through which flows a dense network of capillaries where the gas exchange between air and blood takes place.
The epithelium that lines the alveoli is made up of two cell types, called type I and type II pneumocytes (Figure 2). Type I pneumocytes are flat cells that cover most of the alveolar surface and gases can easily pass through their cytoplasm. Type II pneumocytes occur alone or in small groups among type I pneumocytes, mainly on the edges, where they join the alveolar septa. Their morphology varies from rounded to cubic, with few microvilli on their free surface. Their cytoplasm contains organelles, called lamellar bodies or cytosomes. Cytosomes excrete a surfactant substance on the alveolar surface which mixes with water molecules, reducing their cohesion, thus reducing the surface tension of the alveolar liquid, preventing the lung from collapsing and promoting the need for a smaller inspiratory effort needed to easily to fill the alveoli with air. These cells have the capacity to divide and differentiate into type I pneumocytes, being the main source of cell renewal The center of the alveolar wall is occupied by the interstitium, which is connective in nature and supports a wide capillary network anastomoses.
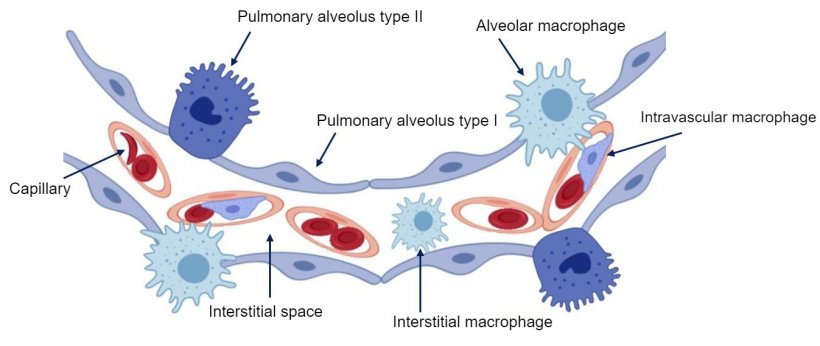
Figure 2: Diagram of the the pulmonary alveolar wall.
Lung macrophages are another important lung defense mechanism. In addition to alveolar macrophages, located in the alveolar lumen, we can also find intravascular pulmonary macrophages, located inside the capillaries, and interstitial macrophages, found in the interstitium of the septa (Figure 2). The former defend the alveolar lumen by phagocytizing microorganisms and inhaled particles, while the latter are responsible for eliminating particles or pathogens that reach the lung via the bloodstream. The population of lung macrophages is the main producer of inflammation mediators, such as cytokines and chemokines, which can also be secreted by other cell types. The main function of these molecules is to attract inflammatory cells such as neutrophils and lymphocytes to the site of injury, thus serving as a "communication system" between the cells involved in the inflammatory process. Viruses such as porcine reproductive and respiratory syndrome (PRRSV) whose main cell targets are alveolar macrophages (Figure 3), alter their basic functions (phagocytosis, antigen presentation and cytokine production) and cause them to eventually die by necrosis and/or apoptosis, which modulates the immune response, producing a delay in the emergence of an effective adaptive response, thus favoring the emergence of secondary infections.
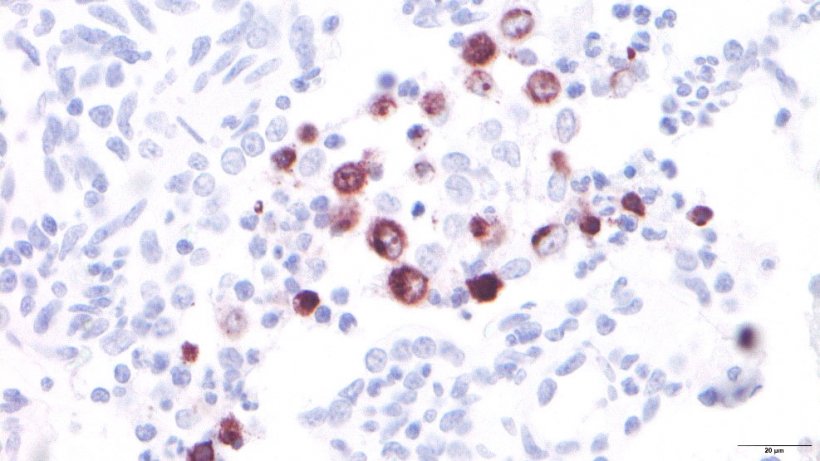
Figure 3: PRRSV-infected alveolar macrophages.
We must never forget that the alveolus, the place where the respiratory exchange takes place, has contact with the exterior, so the integrity of the mucociliary apparatus and the functionality of the macrophages is fundamental to maintain the homeostasis of the respiratory function.




